

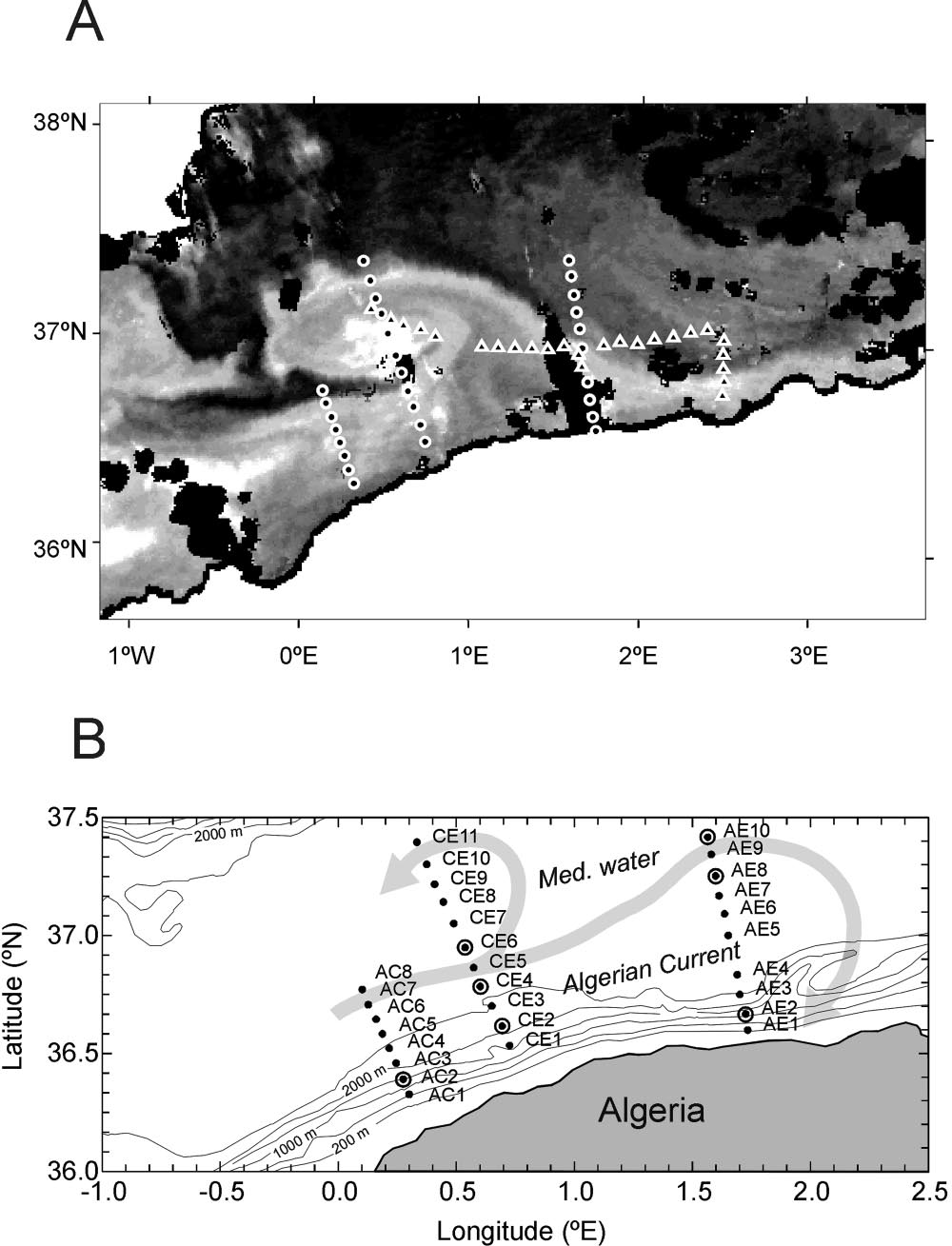
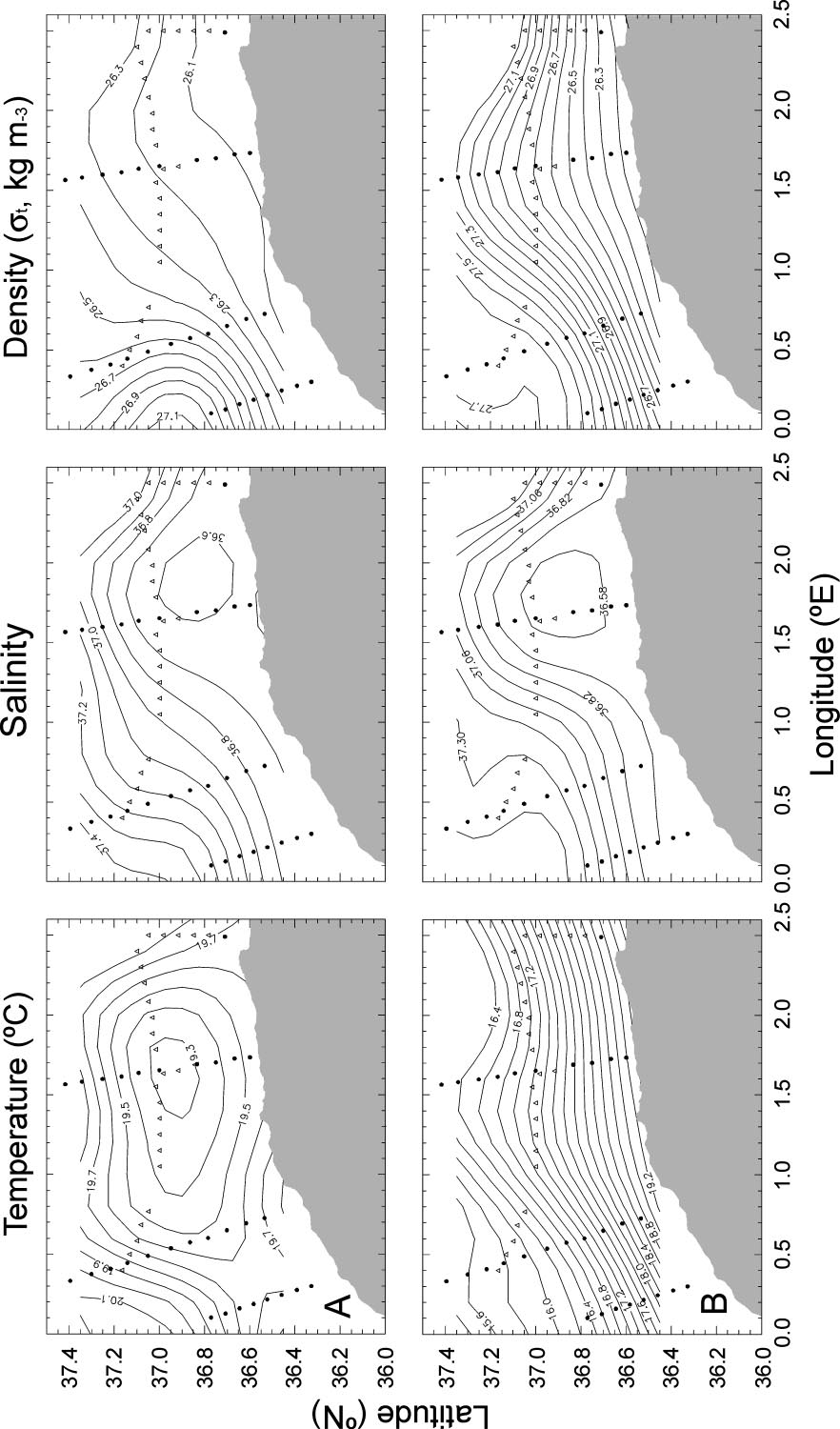
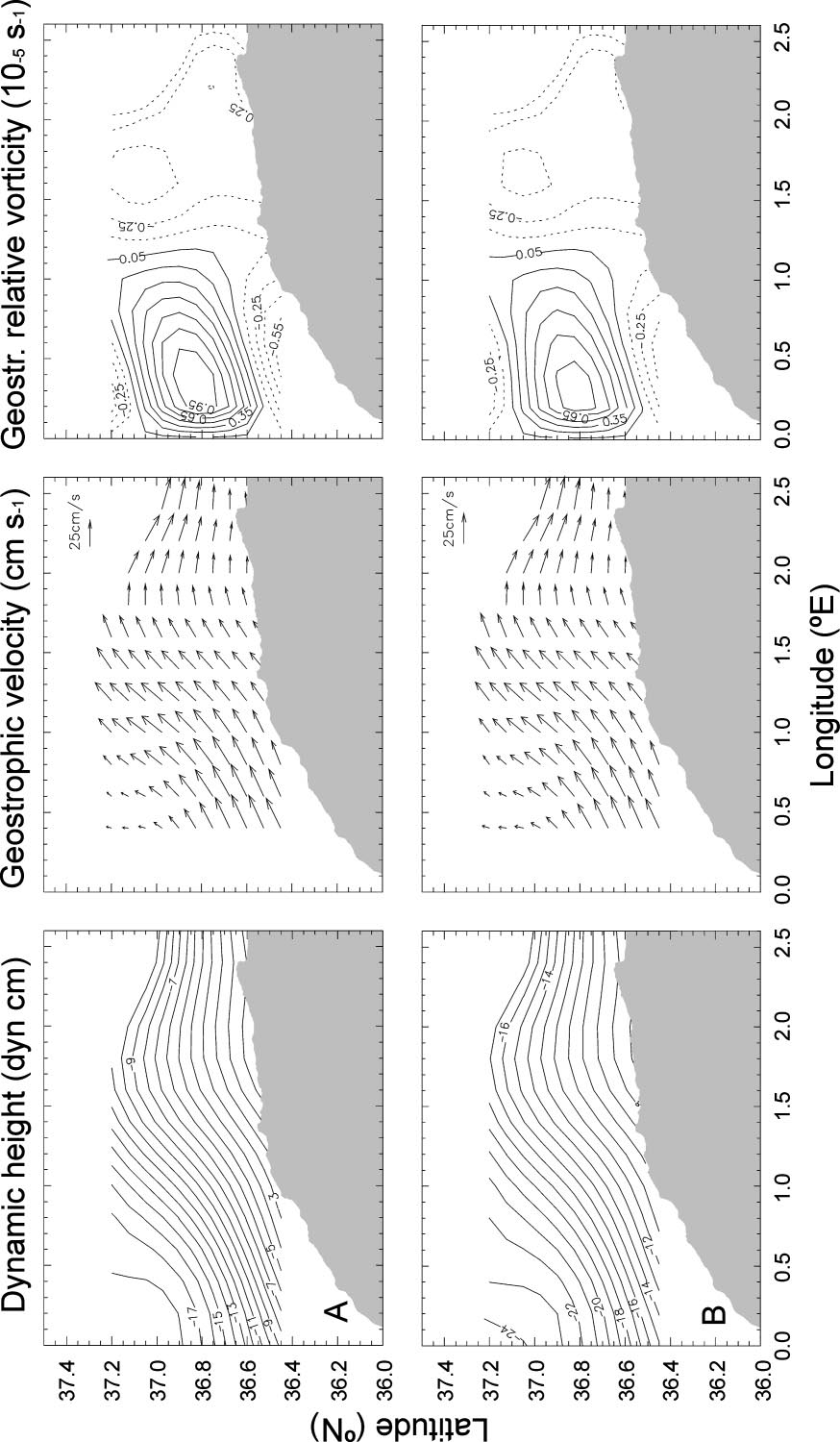

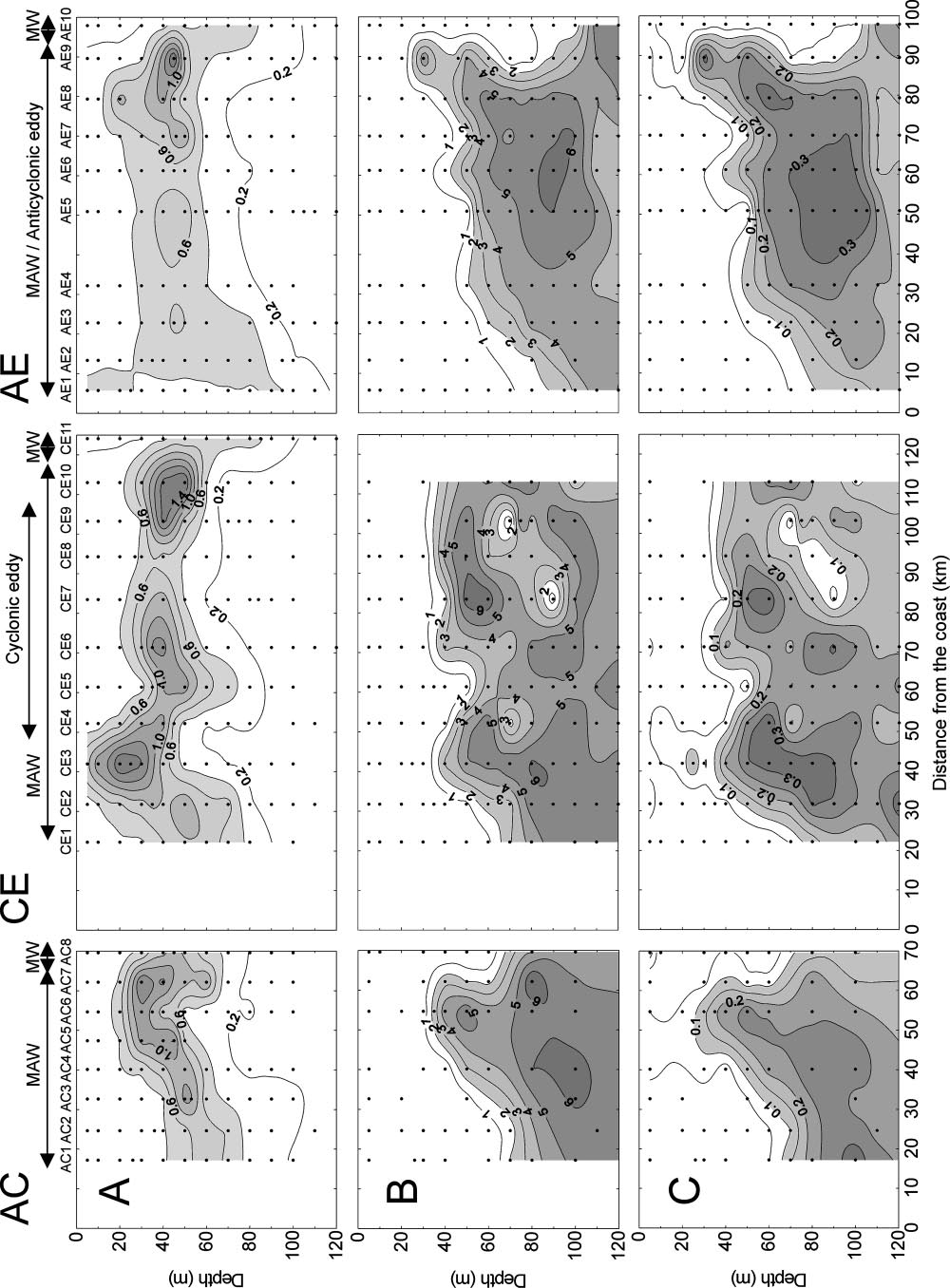
Microsoft word - second edition sept 2012.doc
Directors of Pharmacy NHS Scotland Formulary of Pre-Labelled Medicines for Unscheduled Care Second Edition September 2012 FOREWORD This Second Edition of the NHS Scotland Formulary of Pre-Labelled Medicines for Unscheduled Care has built upon the principles of rationalisation outlined in the 1st Edition and has widened the scope of the document to produce a Formulary containing those pre-labelled medicines that are required in unscheduled care settings. In addition, extensive information on anti-infective prescribing and preparations has been included based on best practice outlined with current HPA Guidance. This document is a valuable resource for NHS Scotland and should help to standardise practice across Health Boards. The Working Group is to be congratulated on the work undertaken to review and up-date this guidance and is asked to continue this valuable work in the future. The Directors of Pharmacy will support the local implementation of this revised Formulary and commend this document to you. Evelyn McPhail Chair Directors of Pharmacy NHS Scotland