Doi:10.1016/j.tips.2007.02.006
TRENDS in Pharmacological Sciences
The Akt–GSK-3 signaling cascade inthe actions of dopamine
Jean-Martin Beaulieu, Raul R. Gainetdinov and Marc G. Caron
Department of Cell Biology, Duke University Medical Center, Durham, NC 27710, USA
Drugs that act on dopamine neurotransmission are
in a diminution of PKA activity D2-class
important tools for the management of multiple neurop-
receptors also modulate intracellular Ca2+ levels by acting
sychiatric disorders. Classically, dopamine receptors have
on ion channels or by triggering the release of intracellular
been shown to regulate cAMP–PKA (protein kinase A) and
Ca2+ stores . Downstream from PKA, dopamine- and
Ca2+ pathways through G-protein-mediated signaling.
cAMP-regulated phosphoprotein with molecular weight 32
However, it has become apparent that, in addition to this
(DARPP-32) has important functions in regulating the
canonical action, D2-class dopamine receptors can func-
efficacy of dopamine receptor signaling and its integration
tion through a protein kinase B (Akt)–GSK-3 (glycogen
with other signaling modalities . Furthermore, extra-
synthase kinase 3) signaling cascade. This novel signaling
cellular-signal-regulated kinase (ERK) has also been ident-
mode involves the multifunctional scaffolding protein b-
ified as an important mediator of cAMP signaling that is
arrestin 2, which has a role in G-protein-coupled receptor
involved in the development of acute and chronic responses
(GPCR) desensitization. In this article, we provide an
to dopaminergic drugs
overview of how this dual function of components of
Recent investigations have shown that, apart from their
the GPCR desensitization machinery relates to dopa-
canonical actions on G-protein-mediated signaling and the
mine-receptor-mediated responses and we summarize
regulation of the cAMP–PKA pathway, dopamine recep-
recent insights into the relevance of the Akt–GSK-3 sig-
tors exert their effects in vivo through cAMP-independent
naling cascade for the expression of dopamine-associated
mechanisms. This new mode of dopamine receptor signal-
behaviors and the actions of dopaminergic drugs.
ing involves proteins that have classically been implicatedin GPCR desensitization Moreover, cAMP-indepen-
dent dopamine receptor signaling displays different kinetic
properties and might serve as an integrator of dopamine
G-protein-coupled receptors (GPCRs) mediating slow
receptor signaling and signaling events that emanate from
neurotransmission Dopamine has an important role
other neurotransmitter receptors.
in the modulation of fast glutamate- and GABA-mediated
In this article, we provide an overview of recent
neurotransmission and is involved in crucial brain func-
advances in the characterization of the dual functions of
tions such as movement, emotion, reward and affect .
dopamine-receptor-desensitizing mechanisms that act at
Consequently, drugs acting on dopamine neurotrans-
the same time as terminators and mediators of different
mission have become widely used tools for the manage-
modalities of dopamine receptor signaling. We also high-
ment of multiple neuropsychiatric disorders, including
light the potential relevance of the novel signaling mech-
schizophrenia, mood disorders, Parkinson's disease, atten-
anism that involves the protein kinase B (Akt)–GSK-3
tion deficit hyperactivity disorder (ADHD) and Tourette
(glycogen synthase kinase 3) pathway () for the
syndrome . A major dopamine-containing region of
expression of dopamine-associated behaviors and the
the brain, the nigrostriatal system, comprises dopamine-
action of dopaminergic drugs.
containing neurons that arise from the substantia nigraand the ventral tegmental area, which project to GABA-
Receptor desensitization mechanisms
containing medium spiny neurons in the caudate putamen
Following receptor stimulation, GPCR signaling is rapidly
and nucleus accumbens (striatum) .
inactivated by a series of mechanisms that results in recep-
Classically, the functions of dopamine receptors have
tor desensitization, internalization and termination of sig-
been associated with the regulation of cAMP–PKA (protein
naling. GPCR activation leads to the rapid phosphorylation
kinase A) through G-protein-mediated signaling . Two
of the receptors by members of a family of GPCR kinases
classes of GPCR mediate dopamine functions. The D1 class
(GRKs) . The phosphorylation of receptors by GRKs
of receptors (D1 and D5 receptors) couple mostly to Gas and
leads to the recruitment of scaffolding proteins termed
stimulate the production of the second messenger cAMP
arrestins, resulting in the uncoupling of the receptors from
and the activity of PKA. By contrast, the D2 class of
G proteins . Two arrestins, b-arrestin 1
receptors (D2, D3 and D4 receptors) couple to Gai/o to
and b-arrestin 2, are expressed in most mammalian tissues,
regulate the production of cAMP negatively, thus resulting
including the brain, whereas two other proteins, the visualarrestins, are expressed specifically in retinal cones androds . The interaction of arrestins with GPCRs is fol-
Corresponding author: Caron, M.G. ().
Available online 8 March 2007.
lowed by the recruitment of an endocytic complex, which
0165-6147/$ – see front matter ß 2007 Elsevier Ltd. All rights reserved. doi:
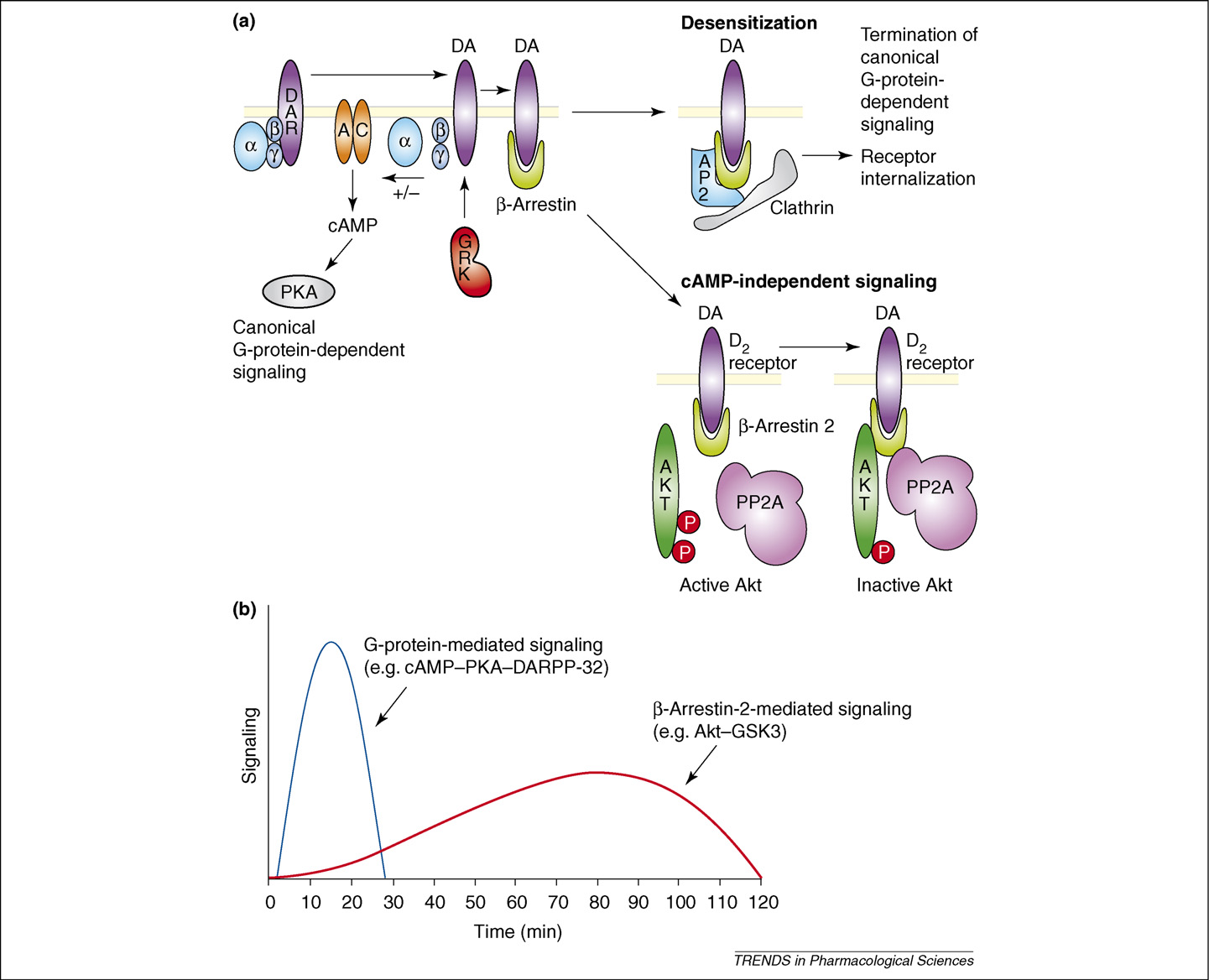
TRENDS in Pharmacological Sciences
Figure 1. Dual role of b-arrestins in GPCR-mediated slow synaptic transmission. (a) The stimulation of dopamine (DA) receptors (DARs) leads to an initial change in receptorconformation that mediates the activation of G proteins, leading to activation (Gas) or inhibition (Gai/o) of adenylyl cyclase and modulation of cAMP-dependent PKA(canonical G-protein-dependent signaling). Receptor phosphorylation by GRKs and the recruitment of b-arrestins then ensue. The recruitment of b-arrestins to receptorsresults in two distinct processes: the termination of G-protein-dependent signaling and the formation of an internalization complex comprising b-arrestin 1 and/or b-arrestin2, adaptor protein 2 (AP2), clathrin and other intermediates. Formation of the internalization complex leads to receptor internalization through clathrin-mediatedendocytosis. The recruitment of b-arrestin 2 following the activation of D2-class receptors also results in the formation of a signaling complex that comprises at least b-arrestin 2, PP2A and Akt. The formation of this complex results in the deactivation of Akt by PP2A and the subsequent stimulation of GSK-3-mediated signaling. (b) Thedifferent kinetics of G-protein-mediated and b-arrestin-2-mediated dopamine receptor signaling following the administration of amphetamine. Shown are the two waves ofsignaling responses involved in slow dopamine synaptic transmission. In the first wave of responses, G-protein-mediated signaling induces a rapid and transient change inthe phosphorylation of direct or indirect PKA targets such as DARPP-32, cAMP-response-element-binding protein (CREB) and ERK. In parallel to these events, a second waveof signaling, mediated by the Akt–b-arrestin-2–PP2A complex, results in a more progressive and longer-lasting response.
leads to the arrestin-dependent internalization of receptors,
to three classes: GRK1-like, GRK2-like and GRK4-like
principally in clathrin-coated pits (a).
. Whereas GRK1 (rhodopsin kinase) and GRK7 (iodop-
Although arrestins were originally identified as key mol-
sin kinase) are found primarily in the retina and are
ecules that control GPCR desensitization, recent evidence
involved in the termination of phototransduction by opsins,
indicates that arrestins and, potentially, GRKs also promote
GRK2, GRK3, GRK4, GRK5 and GRK6 are widely
novel G-protein-independent signaling events that, in
expressed in the body, including the brain. Thus, mice
the case of dopamine receptors, do not involve a regulation
deficient in GRK3 (Adrbk2) GRK4 (Gprk21) ,
of cAMP-mediated signaling
GRK5 (Gprk5) or GRK6 (Gprk6) have beenexamined for their behavioral responses to dopaminergic
Role of GRKs in the regulation of dopamine functions
drugs. Mice that lack GRKs are normal in most tests {with
Studies of dopamine receptor function in mice lacking
the exception of the embryonically lethal GRK2 (Adrbk1)-
different GRKs have, in most cases, supported a role for
knockout mice until challenged with an appropriate
these kinases in both GPCR desensitization and signaling
agonist Because the GRK–b-arrestin system has dual
The seven GRKs in mammals (GRK1–GRK7) belong
roles, both suppressing G-protein-mediated signaling and
TRENDS in Pharmacological Sciences
Box 1. Akt and GSK-3 signaling
hydrobromide] . Thus, although it seems unlikely that
Akt is a serine/threonine kinase that is regulated through phospha-
GRK3 is directly involved in the desensitization of dopamine
tidylinositol-3-kinase-mediated signaling . The regulation of Akt
receptors, it is possible that this kinase is involved in
has been associated with the action of insulin, insulin-related
G-protein-independent dopamine receptor signaling.
peptides [e.g. insulin-like growth factor (IGF)] and neurotrophins[e.g. nerve growth factor, brain-derived neurotrophic factor (BDNF)and neurotrophin 3], which exert their biological function by
Role of b-arrestin 2 in cAMP-independent dopamine
stimulating RTK (see in the main text). The kinases
receptor signaling and function
GSK-3a and GSK-3b are constitutively active and can be inactivated
Several lines of evidence indicate that, in addition to their
through the phosphorylation of the single serine residues serine 21
canonical action on G proteins, GPCRs can also activate
(GSK-3a) and serine 9 (GSK-3b), which are located in theirregulatory N-terminal domains, by Akt and other kinases . Akt
signaling through molecules that are classically involved
inhibits GSK-3a and GSK-3b in response to multiple hormones and
in the regulation of GPCR desensitization (a).
growth factors, including insulin, IGF and BDNF .
Evidence from heterologous cellular systems demonstratesthat b-arrestins can act as G-protein-independent medi-ators of signaling by scaffolding other proteins such as
promoting non-G-protein signaling ), the loss of
kinases and their substrates . However, the
GRKs can have opposing effects on physiological function:
implication of b-arrestin-mediated signaling in biologically
either promoting unregulated (supersensitive) receptor
relevant processes such as slow synaptic transmission
responses because of deficient desensitization or decreas-
has remained largely unexplored. Interestingly, both
ing responses because of a reduced activation of arrestin-
b-arrestin 1 (Arrb1)-knockout and b-arrestin 2 (Arrb2)-
mediated non-G-protein signaling pathways.
knockout mice display reduced behavioral responses to
In mice that are heterozygous for GRK2 deletion,
the dopamine receptor agonist apomorphine Mice
locomotor responses to the psychostimulants cocaine and
lacking b-arrestin 1 also have a reduced responsiveness to
amphetamine or to the direct dopamine receptor agonist
cocaine, whereas b-arrestin-2-knockout mice show blunted
apomorphine are essentially normal, although certain
locomotor responses to the dopamine-dependent actions of
doses of cocaine induce slightly enhanced locomotor acti-
amphetamine and morphine . Furthermore, in
vation Thus, the impact of a partial loss of GRK2 on
mice lacking the dopamine transporter [DAT (Slc6a3)-
dopamine-receptor-mediated responses seems to be lim-
knockout mice], the locomotor hyperactivity phenotype
ited, although it is possible that a more pronounced level of
associated with enhanced dopaminergic tone can be antag-
GRK2 deficiency is necessary to reveal the potential invol-
onized by a concomitant lack of b-arrestin 2 . These
vement of this kinase in dopamine receptor regulation .
observations indicate that one or many different types of b-
Further studies involving brain-specific GRK2-knockout
arrestin-containing protein complexes might mediate
mice will be necessary to address this issue.
dopamine receptor signaling. We have recently identified
GRK6 is the most prominent GRK in the striatum. In
one of these mechanisms and shown that b-arrestin 2 is a
mice, GRK6 is expressed at high levels in the major striatal
signaling intermediate implicated in the cAMP-indepen-
neuronal populations, including GABA-containing med-
dent regulation of Akt and GSK-3 by dopamine
ium spiny neurons and acetylcholine-containing inter-
neurons GRK6-knockout mice show significant
Investigations of cell signaling in response to persistently
supersensitivity to cocaine, amphetamine, morphine and
the endogenous ‘trace amine' b-phenylethylamine, which
reduction of Akt phosphorylation and activity in the stria-
all induce psychomotor activation through the activation of
tum of DAT-knockout mice The inactivation of Akt in
dopamine neurotransmission Detailed investigations
these mice results in concomitant activation of the sub-
of these effects revealed that GRK6-knockout mice have an
strates GSK-3a and GSK-3b which are inhibited by
enhanced coupling of striatal D2-like receptors to G
Akt (Further characterization of these signaling
proteins, an increased affinity of D2, but not D1, receptors
responses using dopamine depletion or dopamine
and enhanced locomotor responses to direct dopamine
receptor antagonists in DAT-knockout mice showed that
receptor agonists . Overall, these observations demon-
Akt, GSK-3a and GSK-3b are regulated by D2-class recep-
strate that cAMP-mediated signaling by postsynaptic D2-
tors ). Furthermore, the D2-class receptor
class receptors in the striatum is regulated by GRK6 and
antagonist and antipsychotic haloperidol leads to enhanced
that D2 receptor desensitization is reduced in the absence
Akt phosphorylation and to GSK-3 inhibition in non-trans-
genic animals Administration of amphetamine or the
Finally, locomotor responses to cocaine are not altered
nonselective dopamine receptor agonist apomorphine to
in GRK4-knockout mice, which is consistent with the
non-transgenic mice also results in an inhibition of Akt
limited expression of GRK4 in the brain . Similarly,
activity, thus confirming the regulation of the Akt–GSK-3
there are no differences in the effects of cocaine and apo-
pathway by dopamine . Interestingly, the regulation of
morphine in GRK5-knockout mice, indicating that GRK5 is
this pathway by dopaminergic drugs shows different
unlikely to be involved in the regulation of dopamine recep-
kinetics compared with those of canonical
tors However, GRK3-knockout mice have normal
cAMP-mediated events and neither Akt nor GSK-
basal locomotor activity but demonstrate significantly
3 is affected by direct modulation of cAMP levels in the
reduced locomotor responses to cocaine, apomorphine
striatum, indicating that the Akt–GSK-3 pathway is not
and the D1 receptor agonist SKF81297 [R-(+)-6-chloro-7,
controlled by this second messenger By contrast, when
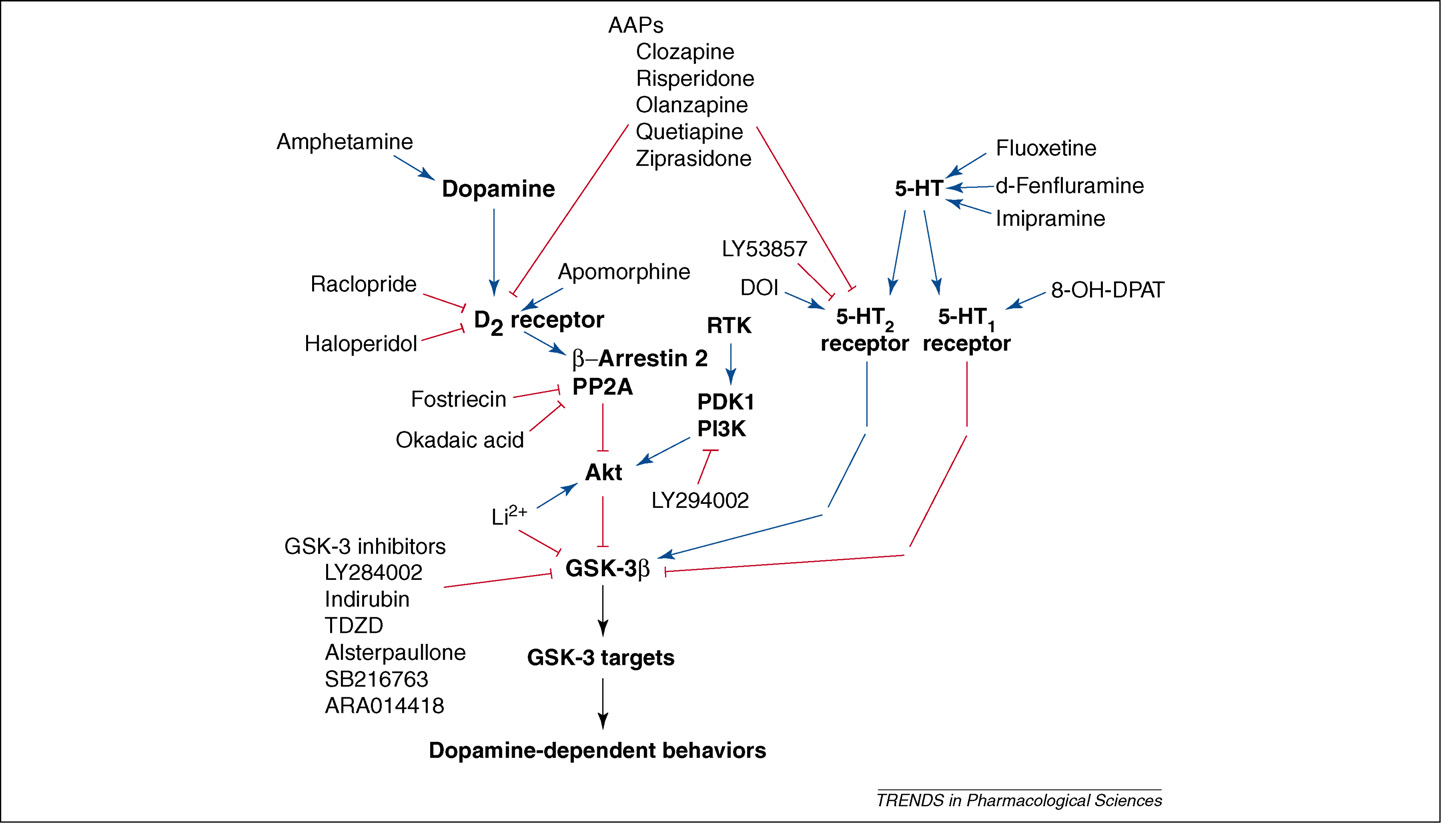
TRENDS in Pharmacological Sciences
Figure 2. Regulation of Akt and GSK-3 by drugs affecting monoamine systems and related signaling events. Behavioral changes in dopaminergic responses have beenreported in Akt1-knockout b-arrestin-2-knockout and GSK-3b heterozygote mice. The GSK-3 inhibitors shown antagonize dopamine-dependent behaviors. Lithium (Li2+) both antagonizes dopamine-associated behaviors and enhances Akt phosphorylation . All other drugs listed are included on the basis of theirreported action on Akt or GSK-3 phosphorylation in the mouse striatum. Blue arrows denote activation, red arrows denote inhibition, black arrows denote effects that canbe either activatory or inhibitory depending on specific substrates. Abbreviations: AAPs, atypical antipsychotics; RTK, receptor tyrosine kinase.
b-arrestin 2 Further investigations of the mechanism
amphetamine nor apomorphine reduces Akt phosphoryl-
by which b-arrestin 2 regulates Akt in response to dopamine
ation Furthermore, mice lacking both b-arrestin 2
showed that inhibitors of protein phosphatase 2A (PP2A)
and DAT show no inhibitory action of elevated levels of
prevent the inhibition of Akt by dopamine, whereas the
extracellular dopamine on Akt phosphorylation, demon-
stimulation of D2-class receptors causes the formation of
strating that dopamine receptors regulate Akt through
a protein complex that comprises at least Akt, b-arrestin 2and PP2A The formation of this complex facilitates thedephosphorylation and deactivation of Akt by PP2A in
Box 2. Two modalities of slow synaptic transmission
response to dopamine and results in the activation of
In cultured fibroblasts, b-arrestin-mediated signaling has a slower
onset and a more prolonged duration than does GPCR signaling
Likewise, the b-arrestin-2-dependent inhibition of Akt bydopamine in the mouse striatum displays a slower but more
b-arrestin-2-knockout mice, multiple lines of evidence
persistent effect than do signaling events that are regulated by the
support the involvement of the b-arrestin-2–Akt–GSK-3
cAMP–PKA pathway (see in the main text).
pathway in the regulation of dopamine-associated beha-
The cAMP-dependent phosphorylation of ERK2 and DARPP-32
viors. For example, GSK-3 inhibitors can reduce locomotor
peaks and subsides within the first 30 min after the administration of
hyperactivity both in DAT-knockout mice and in amphet-
dopaminergic drugs such as amphetamine and cocaine . Bycontrast, the inhibition of Akt by amphetamine develops progres-
amine-treated wild-type animals Confirmation of
sively during the first 30–60 min of drug action and persists over the
these pharmacological observations was also obtained
duration of the drug behavioral effect .
using genetically engineered animals. GSK-3b (Gsk3b)-
This indicates that the regulation and maintenance of certain
knockout mice die during embryogenesis, whereas GSK-
dopamine-associated behaviors, in addition to the action of some
3b heterozygote mice develop normally without overt
dopaminergic drugs, might depend on two complementary wavesof GPCR signaling responses: a first wave of cAMP-mediated
phenotypes . Evaluation of the behavioral actions
responses with a rapid onset and a relatively short duration, and a
caused by amphetamine revealed that GSK-3b heterozy-
second wave of responses characterized by slower onsets and
gote mice are less responsive to this drug over a range of
longer durations, which are dependent on b-arrestin signaling
doses, thus supporting the involvement of GSK-3b in the
functions. Moreover, the reduced behavioral responsiveness to
development of dopamine-associated behaviors Con-
dopaminergic drugs acting on either D1- or D2-class receptorsobserved in b-arrestin-1-knockout and b-arrestin-2-knockout mice
versely, transgenic mice that express a ‘constitutively
indicates that multiple b-arrestin signaling complexes might
active' GSK-3b mutant lacking an inhibitory phosphoryl-
also mediate dopamine receptor signaling and, potentially, the
ation site develop a locomotor hyperactivity phenotype
action of other GPCRs implicated in slow synaptic transmission.
that is reminiscent of DAT-knockout mice . Finally,
TRENDS in Pharmacological Sciences
mice lacking the Akt isoform Akt1 also show enhanced
whereas stimulation of 5-HT1A receptors has the opposite
disruption of sensory motor gating (pre-pulse inhibition) by
effect This action of atypical antipsycho-
amphetamine but not by glutamate NMDA receptor
tics and 5-HT drugs indicates that Akt and GSK-3 might
antagonists The disruption of sensory motor gating
function as signal integrators for dopamine and 5-HT
by amphetamine, which has been used as a behavioral
transmission, and contribute to the action of drugs on
paradigm to model psychosis in rodents, is efficiently
these neurotransmitter systems However, in the
blocked by antipsychotics such as haloperidol that act on
absence of extensive behavioral studies addressing the
D2-class receptors. Furthermore, Akt1-knockout mice also
function of GSK-3 in the regulation of 5-HT function and
show a more pronounced deficit in response to D2-class, but
the action of 5-HT drugs, this integrative function remains
not D1-class, receptor agonists in cognitive tests associated
with prefrontal cortex functions . Therefore, because
Akt and GSK-3 have been associated with the action of
Akt1 is inhibited following the stimulation of D2-class
the mood stabilizer lithium. Lithium is a direct inhibitor of
receptors , the altered behavioral effects caused by
GSK-3 that can also inhibit the activity of this kinase in
amphetamine and D2-class receptor agonists in Akt1-
cells through an indirect mechanism (involving
knockout mice further support the involvement of Akt
Akt activation . Acute and chronic adminis-
inhibition in dopamine-mediated behavioral responses.
tration of lithium inhibits brain GSK-3 activity in mice,as revealed by enhanced regulatory N-terminal domain
Involvement of Akt and GSK-3 in the action of
phosphorylation . Moreover, GSK-3 inhibitors and
psychotropic drugs
reduced GSK-3b expression both reproduce some of the
Typical antipsychotics such as haloperidol are thought to
behavioral actions of lithium in rodents, including its
exert most of their actions by blocking D2-class receptors,
inhibitory action on dopamine-dependent locomotor hyper-
thus supporting a role for dopamine neurotransmission in
activity . Although the mechanism by which
the etiology of schizophrenia. Recent genetic association
lithium regulates Akt and GSK-3 activity is unclear, these
studies have established a link between a deregulation of
observations indicate that a direct or indirect inhibition of
Akt signaling and schizophrenia. Following transmission–
GSK-3 might contribute to the psychopharmacological
disequilibrium tests, a major association of Akt1 haplo-
actions of lithium, at least in part, by inhibiting dopamine
types with schizophrenia has been reported in several
independent cohorts of schizophrenic patients Furthermore, reduced Akt activity or expression levels
Future perspectives
were also shown in the brains of schizophrenic patients
The characterization of the mechanisms by which
b-arrestins and GRKs contribute to dopamine receptor
Thus, because stimulation of striatal D2-class receptors
signaling is at an early stage and it would be naı¨ve to
by dopamine results in an inhibition of Akt it is
believe that the complete palette of molecular responses
possible that a partial loss of function of Akt1 in schizo-
associated with these molecules has been identified. Of
phrenia results in exacerbated responses to D2 receptor
particular interest, the further characterization of GRKs
stimulation that are similar to those observed in Akt1-
and other modulators of b-arrestins in the dopamine sys-
knockout mice. In the same way, classical antipsychotics
tem might enable researchers to determine the functions of
could correct this imbalance by preventing further
these molecules in regulating the positive and negative
reductions of Akt activity by D2-class receptors (
actions of b-arrestins on dopamine receptor signaling. Also
In addition, so-called atypical antipsychotics have
of interest, the D2-receptor-interacting protein spinophilin
recently been shown to either activate Akt or mimic
has been shown to interfere with b-arrestin functions
Akt activity by increasing the phosphorylation of its sub-
by competing with GRK2 for GPCR binding , and GRK2
strates GSK-3a and GSK-3b Atypical antipsychotics
has been shown to regulate Akt in non-neuronal cells
can be distinguished functionally from typical antipsycho-
indicating a potential role of these molecules in the regu-
tics by their reduced affinity and lower specificity for D2
lation of b-arrestin-mediated D2-class receptor signaling.
receptors. Many atypical antipsychotics display a strong
Furthermore, although behavioral studies indicate that
affinity for 5-hydroxytryptamine (5-HT)2A receptors .
b-arrestin 1 also has signaling functions in dopamine
The atypical antipsychotic clozapine enhances Akt–GSK-3
neurotransmission the molecular mechanism(s) of
signaling in cell culture systems Acute or chronic in
these functions has yet to be identified. Interestingly, b-
vivo administration of multiple atypical antipsychotics –
arrestin 1 has recently been shown to function as a nuclear
including risperidone, olanzapine, clozapine, quetiapine
signaling molecule that regulates chromatin structure and
and ziprasidone – results in an inhibition of GSK-3b in
gene expression in cultured fibroblasts . Such
different brain regions Furthermore, drugs that
changes in chromatin organization have also been
affect 5-HT neurotransmission, such as selective 5-HT-
suggested to participate in the development of long-term
reuptake inhibitors, monoamine oxidase inhibitors and
adaptation and addiction to dopaminergic drugs such as
tricyclic antidepressants, amplify the action of atypical
cocaine thus raising the possibility that b-arrestin 1
antipsychotics on GSK-3b Interestingly, increases
has a role in this phenomenon. Finally, the identification of
in GSK-3 activity were recently reported in the prefrontal
a cAMP-independent (b-arrestin-dependent) modality of
cortex of depressed suicide victims . Two 5-HT recep-
dopamine receptor signaling that involves the Akt–GSK-3
tors seem to have antagonistic roles in regulating GSK-3b:
cascade indicates that dopamine receptor functions are
stimulation of 5-HT2A receptors leads to kinase activation,
mediated by multiple mechanisms that collaborate to
TRENDS in Pharmacological Sciences
fine-tune the expression of dopaminergic responses under
21 Peppel, K. et al. (1997) G protein-coupled receptor kinase 3 (GRK3)
different physiological and environmental conditions.
gene disruption leads to loss of odorant receptor desensitization. J.
Biol. Chem. 272, 25425–25428
These mechanisms could be crucial for regulating the
22 Gainetdinov, R.R. et al. (1999) Muscarinic supersensitivity and
expression of distinct dopamine-associated behaviors
impaired receptor desensitization in G protein-coupled receptor
and/or for ensuring the robustness of important phys-
kinase 5-deficient mice. Neuron 24, 1029–1036
iological outcomes. Understanding the complexity of this
23 Gainetdinov, R.R. et al. (2003) Dopaminergic supersensitivity in G
interplay between these modalities of dopamine receptor
protein-coupled receptor kinase 6-deficient mice. Neuron 38, 291–303
signaling could enable the development of novel pharma-
24 Jaber, M. et al. (1996) Essential role of b-adrenergic receptor kinase 1
cological treatments with improved therapeutic actions
in cardiac development and function. Proc. Natl. Acad. Sci. U. S. A. 93,
while avoiding undesired side-effects.
25 Lefkowitz, R.J. and Shenoy, S.K. (2005) Transduction of receptor
signals by b-arrestins. Science 308, 512–517
This work was supported in part by National Institutes of Health grants
26 Bohn, L.M. et al. (2003) Enhanced rewarding properties of morphine,
DA-13511, NS-19576, MH-73853 and MH-40159 (to M.G.C.). M.G.C. is
but not cocaine, in b-arrestin-2 knock-out mice. J. Neurosci. 23, 10265–
the Lattner Foundation National Alliance for Research on Schizophrenia
and Depression (NARSAD) Distinguished Investigator. J-M.B. is the
27 Sotnikova, T.D. et al. (2005) Dopamine-independent locomotor actions
NARSAD Southwest Florida Investigator and the recipient of a
of amphetamines in a novel acute mouse model of Parkinson disease.
fellowship from the Canadian Institutes of Health Research.
PLoS Biol. 3, e271
28 Emamian, E.S. et al. (2004) Convergent evidence for impaired
AKT1–GSK3b signaling in schizophrenia. Nat. Genet. 36, 131–
1 Missale, C. et al. (1998) Dopamine receptors: from structure to function.
Physiol. Rev. 78, 189–225
29 Gould, T.D. et al. (2004) AR-A014418, a selective GSK-3 inhibitor,
2 Greengard, P. (2001) The neurobiology of slow synaptic transmission.
produces antidepressant-like effects in the forced swim test. Int. J.
Science 294, 1024–1030
Neuropsychopharmacol. 7, 387–390
3 Carlsson, A. (2001) A paradigm shift in brain research. Science 294,
30 Hoeflich, K.P. et al. (2000) Requirement for glycogen synthase kinase-
3b in cell survival and NF-kB activation. Nature 406, 86–90
4 Gainetdinov, R.R. et al. (2002) Monoamine transporter pharmacology
31 Prickaerts, J. et al. (2006) Transgenic mice overexpressing glycogen
and mutant mice. Trends Pharmacol. Sci. 23, 367–373
synthase kinase 3b: a putative model of hyperactivity and mania. J.
5 Heimer, L. et al. (1995) Basal ganglia, In The Rat Nervous System
Neurosci. 26, 9022–9029
(2nd edn) (Paxinos, G., ed.), pp. 579–629, Academic Press
32 Lai, W.S. et al. (2006) Akt1 deficiency affects neuronal morphology and
6 Valjent, E. et al. (2006) Role of the ERK pathway in psychostimulant-
predisposes to abnormalities in prefrontal cortex functioning. Proc.
induced locomotor sensitization. BMC Neurosci. 7, 20
Natl. Acad. Sci. U. S. A. 103, 16906–16911
7 Valjent, E. et al. (2005) Regulation of a protein phosphatase cascade
33 Bajestan, S.N. et al. (2006) Association of AKT1 haplotype with the risk
allows convergent dopamine and glutamate signals to activate ERK in
of schizophrenia in Iranian population. Am. J. Med. Genet. B.
the striatum. Proc. Natl. Acad. Sci. U. S. A. 102, 491–496
Neuropsychiatr. Genet. 141, 383–386
8 Beaulieu, J.M. et al. (2006) Paradoxical striatal cellular signaling
34 Schwab, S.G. et al. (2005) Further evidence for association of variants
responses to psychostimulants in hyperactive mice. J. Biol. Chem.
in the AKT1 gene with schizophrenia in a sample of European sib-pair
281, 32072–32080
families. Biol. Psychiatry 58, 446–450
9 Beaulieu, J.M. et al. (2005) An Akt/b-arrestin 2/PP2A signaling
35 Ikeda, M. et al. (2004) Association of AKT1 with schizophrenia
complex mediates dopaminergic neurotransmission and behavior.
confirmed in a Japanese population. Biol. Psychiatry 56, 698–700
Cell 122, 261–273
36 Zhao, Z. et al. (2006) Insulin receptor deficits in schizophrenia and in
10 Beaulieu, J.M. et al. (2004) Lithium antagonizes dopamine-dependent
cellular and animal models of insulin receptor dysfunction. Schizophr.
behaviors mediated by an AKT/glycogen synthase kinase 3 signaling
cascade. Proc. Natl. Acad. Sci. U. S. A. 101, 5099–5104
37 Kang, U.G. et al. (2004) The effects of clozapine on the GSK-3-mediated
11 Arriza, J.L. et al. (1992) The G-protein-coupled receptor kinases
signaling pathway. FEBS Lett. 560, 115–119
bARK1 and bARK2 are widely distributed at synapses in rat brain.
38 Li, X. et al. (2007) Regulation of mouse brain glycogen synthase
J. Neurosci. 12, 4045–4055
kinase-3 by atypical antipsychotics. Int. J. Neuropsychopharmacol.
12 Benovic, J.L. et al. (1991) Cloning, expression, and chromosomal
localization of b-adrenergic receptor kinase 2. A new member of the
39 Kapur, S. and Remington, G. (2001) Atypical antipsychotics: new
receptor kinase family. J. Biol. Chem. 266, 14939–14946
directions and new challenges in the treatment of schizophrenia.
13 Gainetdinov, R.R. et al. (2004) Desensitization of G protein-coupled
Annu. Rev. Med. 52, 503–517
receptors and neuronal functions. Annu. Rev. Neurosci. 27, 107–144
40 Alimohamad, H. et al. (2005) Antipsychotics alter the protein
14 Shenoy, S.K. and Lefkowitz, R.J. (2003) Multifaceted roles of b-
expression levels of b-catenin and GSK-3 in the rat medial
arrestins in the regulation of seven-membrane-spanning receptor
prefrontal cortex and striatum. Biol. Psychiatry 57, 533–542
trafficking and signalling. Biochem. J. 375, 503–515
41 Karege, F. et al. (2007) Alteration in kinase activity but not in protein
15 Lohse, M.J. et al. (1990) b-Arrestin: a protein that regulates b-
levels of protein kinase B and glycogen synthase kinase-3b in ventral
adrenergic receptor function. Science 248, 1547–1550
prefrontal cortex of depressed suicide victims. Biol. Psychiatry 61, 240–
16 Ferguson, S.S. et al. (1996) Role of b-arrestin in mediating agonist-
promoted G protein-coupled receptor internalization. Science 271, 363–
42 Li, X. et al. (2004) In vivo regulation of glycogen synthase kinase-3b
(GSK3b) by serotonergic activity in mouse brain. Neuropsychophar-
17 Laporte, S.A. et al. (2002) b-Arrestin/AP-2 interaction in G protein-
macology 29, 1426–1431
coupled receptor internalization: identification of a b-arrestin binging
43 Beaulieu, J.M. (2007) Not only lithium: regulation of glycogen synthase
site in b2-adaptin. J. Biol. Chem. 277, 9247–9254
18 Luttrell, L.M. et al. (2001) Activation and targeting of extracellular
Neuropsychopharmacol. 10, 3–6
signal-regulated kinases by b-arrestin scaffolds. Proc. Natl. Acad. Sci.
44 Chalecka-Franaszek, E. and Chuang, D.M. (1999) Lithium activates
U. S. A. 98, 2449–2454
the serine/threonine kinase Akt-1 and suppresses glutamate-induced
19 Beaulieu, J.M. and Caron, M.G. (2005) b-Arrestin goes nuclear. Cell
inhibition of Akt-1 activity in neurons. Proc. Natl. Acad. Sci. U. S. A. 96,
20 Shenoy, S.K. et al. (2006) b-Arrestin-dependent, G protein-independent
45 Zhang, F. et al. (2003) Inhibitory phosphorylation of glycogen synthase
ERK1/2 activation by the b2 adrenergic receptor. J. Biol. Chem. 281,
kinase-3 (GSK-3) in response to lithium. Evidence for autoregulation of
GSK-3. J. Biol. Chem. 278, 33067–33077
TRENDS in Pharmacological Sciences
46 Klein, P.S. and Melton, D.A. (1996) A molecular mechanism for the
54 Chao, J. and Nestler, E.J. (2004) Molecular neurobiology of drug
effect of lithium on development. Proc. Natl. Acad. Sci. U. S. A. 93,
addiction. Annu. Rev. Med. 55, 113–132
55 Stelling, J. et al. (2004) Robustness of cellular functions. Cell 118, 675–
47 Phiel, C.J. and Klein, P.S. (2001) Molecular targets of lithium action.
Annu. Rev. Pharmacol. Toxicol. 41, 789–813
56 Scheid, M.P. and Woodgett, J.R. (2001) PKB/AKT: functional
48 De Sarno, P. et al. (2002) Regulation of Akt and glycogen synthase
insights from genetic models. Nat. Rev. Mol. Cell Biol. 2, 760–
Neuropharmacology 43, 1158–1164
57 Chen, M.J. and Russo-Neustadt, A.A. (2005) Exercise activates the
phosphatidylinositol 3-kinase pathway. Brain Res. Mol. Brain Res.
haploinsufficiency mimics the behavioral and molecular effects of
lithium. J. Neurosci. 24, 6791–6798
58 Frame, S. and Cohen, P. (2001) GSK3 takes centre stage more than 20
50 Smith, F.D. et al. (1999) Association of the D2 dopamine receptor third
years after its discovery. Biochem. J. 359, 1–16
cytoplasmic loop with spinophilin, a protein phosphatase-1-interacting
59 Ahn, S. et al. (2004) Differential kinetic and spatial patterns of b-
protein. J. Biol. Chem. 274, 19894–19900
arrestin and G protein-mediated ERK activation by the angiotensin II
51 Wang, Q. et al. (2004) Spinophilin blocks arrestin actions in vitro and in
receptor. J. Biol. Chem. 279, 35518–35525
vivo at G protein-coupled receptors. Science 304, 1940–1944
60 Valjent, E. et al. (2000) Involvement of the extracellular signal-
52 Liu, S. et al. (2005) A crucial role for GRK2 in regulation of endothelial
regulated kinase cascade for cocaine-rewarding properties. J.
cell nitric oxide synthase function in portal hypertension. Nat. Med. 11,
Neurosci. 20, 8701–8709
61 Svenningsson, P. et al. (2003) Diverse psychotomimetics act through a
53 Kang, J. et al. (2005) A nuclear function of b-arrestin 1 in GPCR
common signaling pathway. Science 302, 1412–1415
signaling: regulation of histone acetylation and gene transcription.
62 Beaulieu, J.M. et al. (2007) Regulation of Akt signaling by D2 and D3
Cell 123, 833–847
dopamine receptors in vivo. J. Neurosci. 27, 881–885
Elsevier.com – linking scientists to new research and thinking
Designed for scientists' information needs, Elsevier.com is powered by the latest technology with
customer-focused navigation and an intuitive architecture for an improved user experience and
The easy-to-use navigational tools and structure connect scientists with vital information – all
from one entry point. Users can perform rapid and precise searches with our advanced search
functionality, using the FAST technology of Scirus.com, the free science search engine. Users can
define their searches by any number of criteria to pinpoint information and resources. Search by a
specific author or editor, book publication date, subject area – life sciences, health sciences,
physical sciences and social sciences – or by product type. Elsevier's portfolio includes more than
1800 Elsevier journals, 2200 new books every year and a range of innovative electronic products.
In addition, tailored content for authors, editors and librarians provides timely news and updates
on new products and services.
Elsevier is proud to be a partner with the scientific and medical community. Find out more about
our mission and values at Elsevier.com. Discover how we support the scientific, technical and
medical communities worldwide through partnerships with libraries and other publishers, and grant
awards from The Elsevier Foundation.
As a world-leading publisher of scientific, technical and health information, Elsevier is dedicated to
linking researchers and professionals to the best thinking in their fields. We offer the widest and
deepest coverage in a range of media types to enhance cross-pollination of information,
breakthroughs in research and discovery, and the sharing and preservation of knowledge.
Elsevier. Building insights. Breaking boundaries.
Source: http://www.lira.dist.unige.it/IIT_school/files/papers/tips4.pdf
Journal of Power Sources 110 (2002) 321–329 Battery performance models in ADVISOR National Renewable Energy Laboratory, Golden, CO 80401, USA This paper summarizes battery modeling capabilities in ADVISOR—the National Renewable Energy Laboratory's advanced vehicle simulator written in the Matlab/Simulink environment. ADVISOR's Matlab-oriented battery models consist of the following: (1) an internalresistance model, (2) a resistance–capacitance (RC) model, (3) a PNGV capacitance model, (4) a neural network (nnet) lead acid model, and(5) a fundamental lead acid battery model. For the models, the electric schematics (where applicable), thermal models, accuracy, existingdatasets, and sample validation plots are presented. A brief summary of ADVISOR's capabilities for co-simulation with Saber is presented,which links ADVISOR with Saber's lead acid battery model. The models outlined in this paper were presented at the workshop on‘Development of Advanced Battery Engineering Models' in August 2001.# 2002 Elsevier Science B.V. All rights reserved.
Protocolo Nacional Unificado para la Protección y Asistencia Integral a Personas Víctimas de Trata Protocolo Nacional Unificado para la Protección y Asistencia Integral a Personas Víctimas de Trata PROTOCOLO NACIONAL UNIFICADO PARA LA PROTECCIÓN Y ASISTENCIA INTEGRAL A PERSONAS VÍCTIMAS DE TRATA Jefe de Misión OIM EcuadorRogelio Bernal Equipo del Programa Contra la Trata de Personas OIMArgentina SantacruzBelén Rodríguez

