

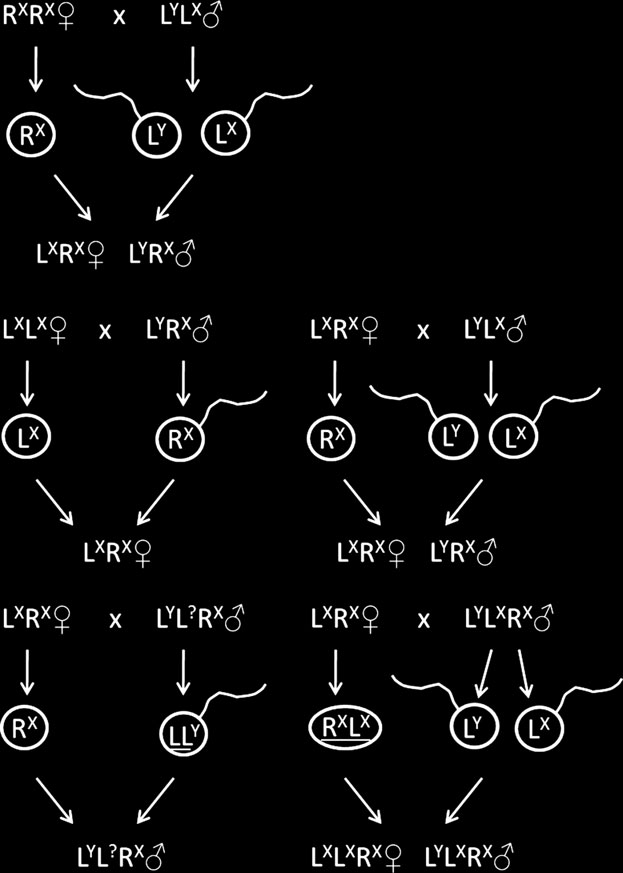
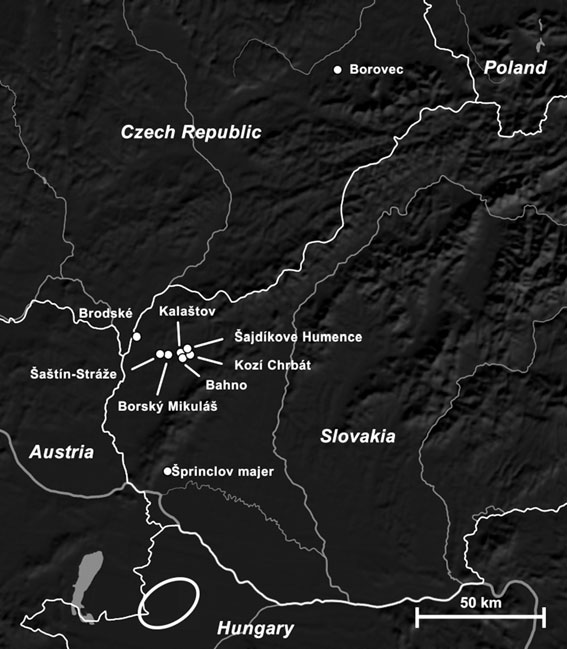
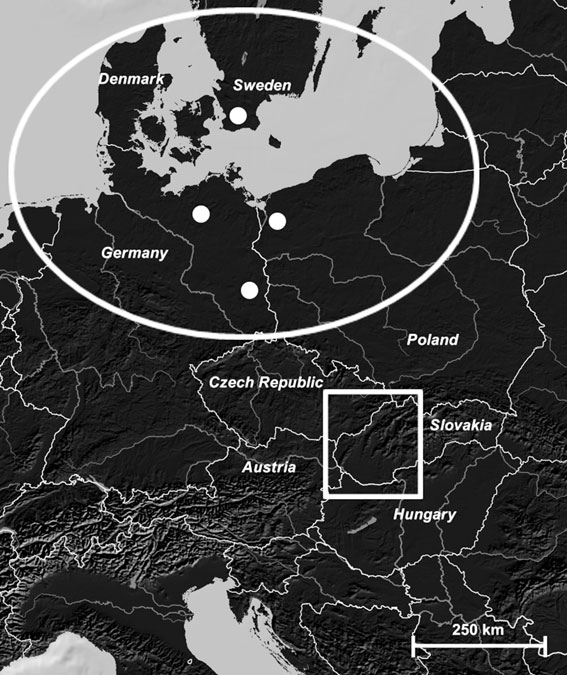
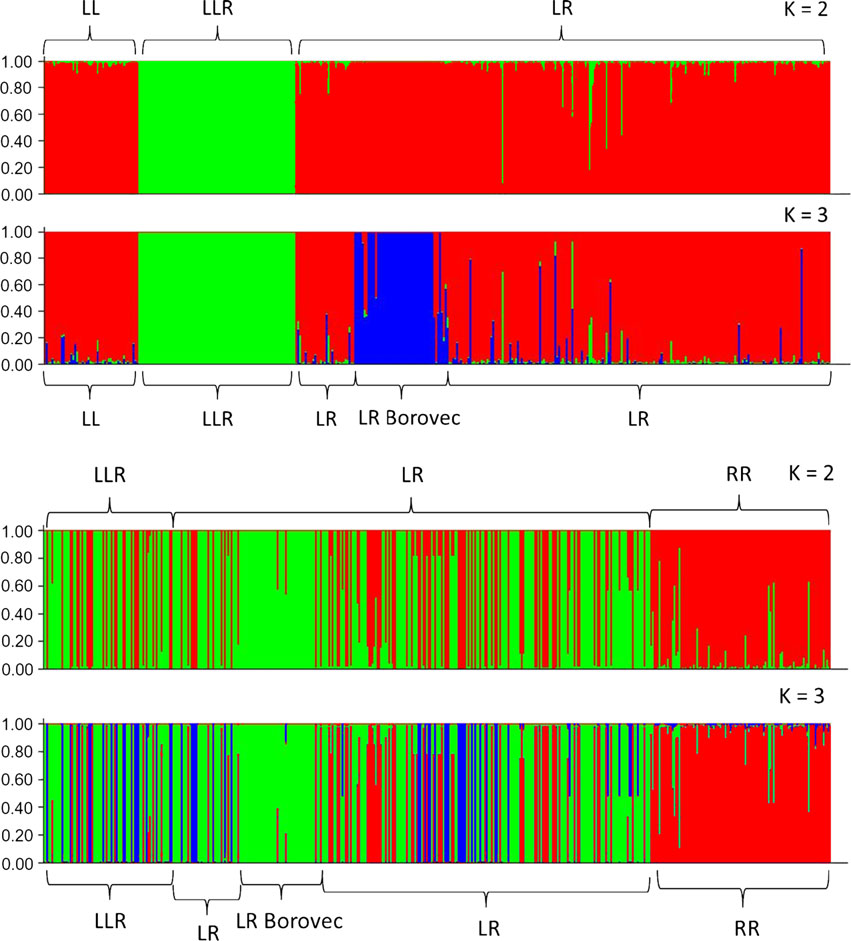
frauenarzt-boeblingen.de
© Schattauer 2011 Diagnostik und Behandlung des Gestationsdiabetes entsprechend den aktuellsten Leitlinien U. Schäfer-Graf Berliner Diabeteszentrum für Schwangere, Klinik für Gynäkologie & Geburtshilfe, St. Joseph Krankenhaus Keywords Definition Gestationsdiabetes, Insulinresistenz in der Gestational diabetes, insulin resistance in